 LOGIN
LOGIN REGISTER
REGISTER.png)


Research Article
A Theoretical Approach Concerning Nutrient Emissions to Inform Monitoring and Management of Mariculture Farms
Leigh A. Kroeger1* Anat Tsemel2, Dan Tchernov2 and Ofira Ayalon1
1Department of Natural Resources and Environmental Management, University of Haifa, Israel
2Morris Kahn Marine Research Station, Department of Marine Biology, Leon H. Charney School of Marine Sciences, University of Haifa, Israel
*Corresponding author: Leigh A. Kroeger, Department of Natural Resources and Environmental Management, University of Haifa, Israel, Email: leigh.kroeger@gmail.com
Citation: Kroeger LA, Tsemel A, Tchernov D and Ayalon O (2019) A Theoretical Approach Concerning Nutrient Emissions to Inform Monitoring and Management of Mariculture Farms. J Aquat Res Mar Sci, 2019: 125-133.
Received Date: 22 January, 2019; Accepted Date: 04 February, 2019; Published Date: 13 March, 2019
Abstract
As demand for production requires stakeholders to consider national marine space to keep up with product demand, it is crucial that the question of environmental impact is prioritized. A multitude of biotic and abiotic challenges face the expansion of the mariculture industry in the ultraoligotrophic waters of the Levant Basin of the eastern Mediterranean Sea (EMS). Several local monitoring efforts have addressed sediment enrichment and water quality, however, estimating the loading of bioavailable nitrogen as a result of farming has not been attempted. In this study, baseline dissolved nutrients and sediment samples were collected in situ at a monoculture farm 3 km offshore (300 t caged biomass of Sparus aurata), and from a control site. Analysis of all samples indicated no significant impact of the fish farms when compared to the control. The ratio of in situ nitrogen (N) to phosphorus (P) was similar to the N:P ratio from a published mesocosm. Thus, the nitrogen excretion rates of gilthead sea bream (Sparus aurata) from this study were utilised to produce a theoretical calculation and waste budget. The calculation was configured to the input of artificial fish feed pellets fed to the mariculture system per day for 700 t caged fish (the governmental limit of production). The input of artificial food pellets will yield 197,252 kg of bioavailable N per year into the water column (540 kg/day). Given this local point source of nutrients, the in situ results and calculation suggest that farming at 700 t should not have significant measurable effects on the marine system and are rapidly assimilated by primary producers. Mixed methods studies combining field collection and published case studies may further strengthen science-based management of the marine space and provide justification in developing long-term monitoring protocols.
Keywords
eastern Mediterranean Sea; mari culture; effluent; particulate matter; dissolved nutrients; nutrient budget; ultraoligotrophic; production limits; Sparus aurata
List of abbreviations
Conductivity-Temperature-Depth (CTD)
Chlorophyll A (Chl-a)
Dissolved Organic Phosphorus (DOP)
Dissolved Organic Nitrogen (DON)
Eastern Mediterranean Sea (EMS)
Fish Aggregating Device (FAD)
Food Conversion Rate (FCR)
Inorganic Carbon (IC)
Organic Matter (OM)
Particulate Organic Matter (POM)
Self-Contained Underwater Breathing Apparatus (SCUBA)
Total Suspended Solids (TSS)
Total Organic Carbon (TOC)
Introduction
Food security is a key concern for humanity in the face of a rise in global population and a quest for new protein sources [UN Sustainable Development Goals, 14; [1]. This has fuelled research and development of the agri-sector, namely of marine sources. As the fastest growing food production sector in recent decades [2], governments are considering developing their national marine space as coastline habitation is increasing [3,4] as well as onshore conflicts of interest with other industries [5]. Mariculture operations make up 43.5% of total aquaculture income and are being practiced in 112 countries as of 2013 [6]. The benefits of offshore operations [> 2 km from the coastline] may include higher oxygen saturation, better meat quality, and lower visual impact and consequences of disease outbreaks [5]. There is also little need to mimic the commodity’s natural environment [7]. However, several studies have explained the potential negative effects of mariculture operations, including nutrient loading, fish escapes, pathogen emergence, and cages as fish aggregating devices [FADs; [8-12]]. In particular, nutrient loading from particulate and soluble fish excretion is well-documented as a potential point and/or diffuse impact surrounding offshore/open water fish cages [13-15]. There are numerous studies describing nutrient footprints and shifts in water and sediment column regimes as a result of mariculture [16-17 and 11-12].
To remain true to the lauded ecosystem approach to sustainable development of industries [5], environmental managers typically employ the concept of assimilative carrying capacity to assist in deciding on limits of mariculture production. Defining a nutrient effluent budget of the cultured species is a high-resolution instrument to achieve, at least in part, a limit to farm production, and requires local groundtruth sampling. This bottom-up research at local scale can clarify shifts and imbalances with negative effects on the ecosystem and is essential to properly managing the resource [18], especially in atypical water bodies such as the eastern Mediterranean Sea [EMS].
The EMS is classed as ultra-oligotrophic [Class III ecosystem; [19]; specifically, the annual average for the Southern Levantine Sea was estimated to be 0.224 g C m−2 d−1 [20].
The marine system is unusual in that the deep waters exhibit a high nitrate to phosphate [N:P] ratio [28:1] and are phosphorus-limited [21]. The ultra-oligotrophic status is caused by a lack of phosphate [surface waters < 10 nM], not preferential removal of nitrogen. Therefore, global or regional nutrient waste budgets for fish farms are not applicable, and these specific parameters must be considered in developing a niche budget for the Levant Basin. Such a budget is essential for understanding trophic dynamics and fluctuations from the bottom-up perspective for environmental managers. However, the sampling of nutrients and detection is difficult in oligotrophic systems due to rapid dilution and assimilation [14-15]. When utilized in combination with an understanding of the farms’ output and geometry, water column characteristics and exchange rates, managers can address these layers of complexity in the management process and mitigate for issues such as increased organic matter on the sediment surface, greater oxygen consumption and/or potential anoxia [22].
Nutrient budgets have been used extensively to understand biogeochemical cycling processes in the eastern Mediterranean, partly because the system is well constrained with marine inputs and outputs occurring through the relatively narrow Straits of Sicily [23,21,24]. Several past studies have focused on the use of Lagrangian
particle tracking to model dissolved and particulate wastes [25-27], however, a detailed budget of nutrient effluent as the result of artificial feed input has not been applied to an offshore site. There are two previous mesocosms published for S. aurata: a) a rate of nitrogen excretion [28] and b) an estimated budget of N and P dissolved and particulate wastes. However, these studies do not have an in situ counterpart.
This study aimed to analyse the water and sediment column nutrient concentrations from a small-scale fish farm located three kilometres offshore Israel to determine if there is a significant difference between the sites. The water column data is then used to develop a theoretical budget of metabolized wastes from harvesting 700 tonnes of Sparus aurata over the government-allocated farm site. It is a micro-scalar study to delineate how much of dissolved and particulate nitrogen is being inputted to the ecosystem, and its biological effect to the natural food chain. This study is also unique in that it combines ground truth data from the site and applies it to a mesocosm study [28]. The results are compared to a large-scale, long-term artificial point source of nutrients, and the government-recommended thresholds of dissolved nutrient concentrations.
Materials and Methods
a. Fish farm site description
The sampling site is located three km offshore from Michmoret, Israel (32º 24' 49.3194" N, 34º 50' 15.6582" E, Figure 1). The farm (Lev Yam Aquaculture Ltd.) consists of four 3,000 m3 tension-leg cages (Reframed, Italy), moored on a flat topography at a depth of 37 meters. They are submerged at a minimum depth of five meters with dimensions being 20 m diameter and 9 m depth. The substrate under the cages is composed of mainly sandy sediments (20% < 63 µ; [29]). Currently, the site produces 300 tons of gilthead sea bream (Sparus aurata) fish per year. The fingerlings are caged at 2-7 grams and harvested at 300-400 g. At the time of the study, there was a fish biomass of ~300 tons and stocking density of 17 kg m3.
b. Sample collection, preservation, and analysis
The sampling of dissolved nutrients was carried out in March 2017 at the fish farm and control sites. During the sampling, the fish were fed at 0900 and 1200 hours but the feeding time varies dependent on the stocking biomass and water temperature. The fish diet changes seasonally (supplementary material A) and the fish are fed 1-2% of their biomass per day. Standard water column measurements using a a CTD (Sea-Bird SBE 911) were taken from the control site and fish farm. A total of five CTD casts were deployed after anchoring approximately 100 m downstream of the fish cages, and two casts were deployed at the control station one km upstream. At both sites, the CTD sampled at five m depth intervals, from 5 to 30 m depth. Two water samples taken from each bottle for nutrients (phosphate, nitrate, ammonium) analysis. The samples were stored in the on-board refrigerator for nutrient analysis the following day. For chlorophyll-a, all water samples were filtered through a Whatman 25 mm GF/A filter and then frozen for subsequent analysis.
The dissolved nutrients were determined using a SEAL AutoAnalyzer AA3 system, according to standard protocols [30]. Phosphate was analysed using the molybdate blue method detected using a LWCC long flow cell, and nitrate was analysed using Cd reduction followed by formation of an azo dye and ammonia by o-phthaladehyde fluorescence dye procedure. Total Dissolved Phosphorus (TDP) was determined as phosphate after addition of persulphate and oxidation using an autoclave for two hours [31]. DOP was calculated as the differene between TDP and Phosphate. Samples for chlorophyll were collected then filtered through pre-weighed Whatman 47 mm GF/F filters (0.7 µm effective pore diameter) and frozen. Chlorophyll was extracted and measured as in the [32] method with a Turner design TD-700 Fluorimeter. For Particulate Organic Matter (POM), and Organic Matter (OM%), sediment samples were collected on board, then filtered through pre-weighed Whatman 47 mm GF/F filters (0.7 µm). The results were determined from a second weighing of the filters after combustion at 450 ºC. Total suspended solids (TSS) was determined after drying, weighing, and concentration calculated.
Sediment samples were taken from a control station (5 km north of fish cages; 32º25’18.8” N, 34º50’22.9” E) and under the fish cages (32º24’48.0” N, 34º50’14.5” E) in March 2017.
|
Figure 1: Selected sites for sampling, the red square in the top left indicates the area of the Mediterranean basin. The fish farm is indicated by a red circle. The control site for water sampling was 1 km south of the cages (32º 24' 30.9996" N, 34º 50' 18.9996" E), and the sediment sampling was 5 km south of the fish cages. The research vessel was anchored 100 m downstream of the fish cages sampling every two hours |
Sediment samples were taken from a control station (5 km north of fish cages; 32º25’18.8” N, 34º50’22.9” E) and under the fish cages (32º24’48.0” N, 34º50’14.5” E) in March 2017.
Four cores were obtained through manual extraction (SCUBA divers) at the fish farm and control sites using Plexiglas push cores of 6.5 cm internal diameter and 20 cm length, to collect samples from the upper 15 cm of sediment. Onboard, the sediment cores were immersed in ice, kept at an upright position, and transported to the laboratory. Two cores were sliced at 0.5 cm depth intervals up to a maximal depth of 10 cm. Each slice was sub-sampled avoiding the edges of the slice and transferred to a sterile polyethylene tube. Sediment samples were freeze-dried over 48 hours in a Heto Lyolab 3000 freeze drier connected to a RZ 5HE Vacuubrand vacuum pump, and 250 mg of dry sediment was taken for the analyses.
Total inorganic and organic carbon [TOC] were determined using a Skalar Primacs SLC analyser. Inorganic carbon [IC] was determined by acidification and purging, while TOC was determined after removing IC fraction using 5% HCl for three hours by combustion at 1050 ºC. Total Phosphorus and Inorganic Phosphorus was determined in these sediment samples using the method of [31].
c. Compilation of estimates from literature
There were several key pieces of data from the published mesocosm studies that were necessary to develop the calculation for bioavailable nitrogen to the marine system (Table 1). First, the researchers had to ensure the mesocosm and in situ data were compatible (see supplementary information B for full comparison). The in situ water sampling was consistent over 8 hours, however the fish were fed twice during the sampling. The ammonia excretion rate study [28] was held over 24 hours, and the fish were fed once in the morning after a starvation period. Thus, the instantaneous excretion rate might differ, although the total amount of effluent remains the same. To guarantee that the current study’s excretion rate was approximate to [28], we compared the in situ N:P ratio to the published estimates S. aurata N:P ratio [33]. Therefore, the relevant data from [28] regarding ammonia excretion was incorporated into the model.
Porter et al. [1987] completed a total ammonia excretion rate and budget for S. aurata in a microcosm for 24 hours (190 litre tanks, 24ºC). The researchers fed their specimens once at 0800 hours and sampled the water every four hours. They determined that for a 90-g fish, approximately 30% of excretion was ammonia-N, and 30% excreted as dissolved organic N (DON). The rest was either faeces (5.6%) or assimilated as fish growth (21.3%). The percentage of sea bream faecal waste from extruded pellets was complemented by other research (4-6%; 34).
Using these percentage estimates, the calculation was then configured to match the Israeli government’s allocated farm production limit (700 t). The researchers used an averaged estimate of artificial fish feed pellets inputted per day (1.5% of stocking biomass). On the day of sampling, the cage biomass was estimated to be 300 tonnes, and the fish were approximately 100 g (wet weight). The fish are, therefore, comparable to [28] 90-g specimen’s excretion rates. The mesocosm’s average water temperature (25 ºC), food input (1-2%), and FCR for S. aurata were comparable to the current cage situation. Thus, the calculation assumes an average of 1.5% of fish food was fed on the day to a 700 tonne stocking biomass, for a 90-g fish. Other assumptions made pre-calculation are as follows:
No percentage of fish food pellets ‘sink’ through the water column and settle on seafloor (i.e. 100% consumption by farmed fish in cages).
- Wild fish species are considered conspecific to Sparidae, thus, excrete at the same rate as S. aurata, as well as share a similar FCR rate
- Food composition of nitrogen equates to [28] estimates of consumed nitrogen per 100 g of Sparus aurata biomass
Table 1. Assembled data estimates and information required for calculation of bioavailable effluent from the fish farm (averages estimated from [28] for 90-g fish)
|
d. Statistical analysis of in situ data
Standard student paired t-tests were used to derive significant differences between the control and fish cages sites, to the 95th percentile. This test was used for differences in DOP, ammonia, and phosphate, organic matter, chl-A, TSS and POM, and TP in sediment. No statistical tests were used for % Total Organic Carbon, or grain size, as there was only one core sample from the farm and control each.
Results
a. Characterisation of the water and sediment column – local and distal effects
In total, 84 samples were collected on board for later analysis of dissolved nutrients. Ammonia-N, DOP and phosphate were chosen for analysis because of their notable importance in primary production. There was an observed increase in dissolved nutrients with time and water column depth, however, there was no significant difference between the control and fish cages (p >
|
Figure 2 a, b, c. Measured concentrations of dissolved organic phosphate (top), ammonium (middle), and phosphate (bottom, nM) at the fish cages (FC) and control sites [see legends]. Concentrations are demarcated by depth (5 m intervals to 35 m), and by cast number (FC1 to FC5). Samples were taken at 2-hour intervals at a single station 100 m from the nexus of the fish cages. Samples below the limit of detection are indicated on the left side of the line in DOP. |
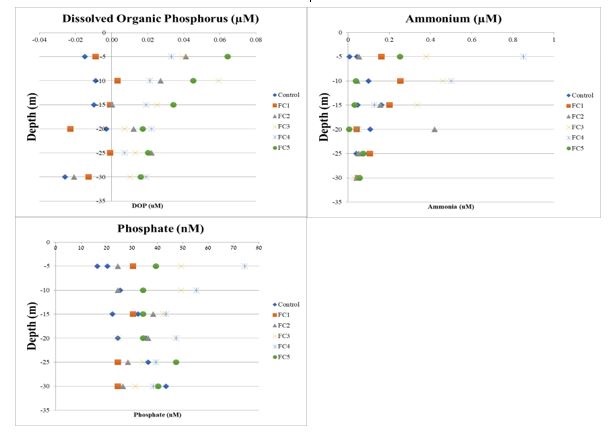
0.05, Figure 2) in terms of dissolved nutrients. There was also no significant difference between chl-1, TSS, or POM samples taken at the control site (1 km upstream) or the fish cage site (p > 0.05 across all depths and temporal variation).
Qualitatively, there was a slight increase in total organic carbon at core depth at both the control and cage sites in the sediment column, which increased with core depth. The % grain size at the fish farms was more uniform than at the control site, with a higher percentage < 63 µ grains with depth at the control. Between the fish farms and control sites, there was no significant difference in total phosphorus within the top 10 cm of sediment (p > 0.05).
b. Calculation of bioavailable nitrogen effluent into the marine system
A full step-by-step explanation of the calculation of bioavailable N excretion and translation to generated primary production can be found in supplementary information C. The simplified results are: the caged fish utilize 21.3% of the inputted N as growth and excrete dissolved N effluent which is considered bioavailable to the phytoplankton community (73.4% of total N input from fish feed or 36,534 mol N; 15). Of the caged particulate effluent ingested on the way down the water column (5.6%), 595.4 mol N is translated as secondary fish consumer growth. The secondary excretion of the wild fish will produce a further 2,048.7 mol of bioavailable N to the system (73.4% of their soluble excreted N). This total amount of N (38,582 mol Nmol N) is translated into kg N per year.
- Discussion
This research delineates the qualitative nature of effluent into the marine ecosystem surrounding an offshore monoculture fish farm. While the field data obtained in this study is direct evidence of the type and even ratio of dissolved nutrients being excreted by the fish into the surrounding water (when compared to the [28] mesocosm), the samples that were collected in this research are not diversified enough to give a good enough estimate of the affect of the fish cages on the environment. To capture all the discharge would require unrealistically intensive sampling in both space and time. As such, the results from water body and sediment core analyses were analysed and then were compared with nutrients waste budgets and excretion rates from the literature to assess compatibility. After confirmation, the missing parameters were supplemented in order to estimate the amount of bioavailable N resulting from an input of fish food into a marine system. Finally, the calculation’s result is discussed in comparison to another artificial source of nutrients at the same depth offshore, and the expansion of the industry. Potentials limits to the expansion are expressed in the ecological context of stated nutrient limits for the state of Israel.
a. In situ measurements and comparison to literature estimates
Field measurements at the fish cages and control sites were analysed to quantify the flux of dissolved and particulate nutrients from the fish cages into the environment. The impact to water and sediment quality, and seasonal flux, is paramount when defining limits to industry expansion; the sampling of pelagic and benthic samples can be equally important to the benthic parameters [35]. Numerous studies have shown that dissolved nutrients from fish cages are assimilated and taken up the food chain almost immediately, which may support why at the 300-t production level, no significant difference was determined [36-37,15].
The dissolved nutrients data were consistent with the [28] laboratory experiments of excretion rates, which finds a peak in ammonia excretion 3-4 hours post-feeding. Furthermore, the N:P ratio was compatible with the ratio from lab experiments (see supplementary information B). It was expected that with a low background of dissolved nutrients, it might make detecting fish cage effluent easier. However, measurements made at the control and cage sites showed no significant increase in nutrients, either due to rapid dilution or uptake, or due to displacement of the plume during sampling. [38] concludes that the nutrient effluent from the Israeli fish farms is rapidly diluted and carried along the continental shelf, with the predominant currents. This is in sharp contrast to typical aquaculture research (i.e. Scottish fjords) and is attributed to the high exposure and ultra-low baseline nutrient concentration of the basin. Although not significant between the farm and control sites, there was a peak in ammonium of 0.9 µM and of phosphate 0.07 µM at five m depth, seven hours after feeding. These are well within the Ministry of Environmental Protection’s limit of 1 mg/L [39], however not within the recommended thresholds of 0.45 and 0.05 µM, respectively, for ‘Spring Season: Alongshore’ [40]. At this production size, it is expected that the nutrient loading may surpass the suggested open water threshold at specific times and depths.
There was a minor increase of chl-a near the bottom of the cages and control site, however not significant; it is noted that other studies have concluded the maximum is a normal occurrence and not necessarily related to the bottom dynamics [41]. All statistical testing determined there was no difference between the control or fish cage site, at the current production level. Therefore, the assumption that all wasted artificial pellets and fish faeces are consumed through the water column by wild fish populations is justified and in line with other regional studies [in situ observations; 41-43]. The results also suggest no difference in grain size or TOC %, nor in Total P (Inorganic and Organic). Thus, any effluent, dissolved or particulate, is taken up into the food chain or rapidly diluted, which creates a ‘hot spot’ food source. Inorganic phosphorus was only slightly higher at the cages than control site.
In past studies, it is commonly accepted that not all artificial pellets are consumed by the farmed commodity, and that the metabolized wastes accumulate locally underneath the cage sites [44-45]. The well-documented farms in offshore waters of the USA, Canada, Norway or UK are generally in sheltered fjords which tend to deposit fine grained, organic rich sediments [46-48]. It is likely that the combination of small production, open water culture and the physical properties of the sandy substrate are the contributing factors that lessen the effluent’s impact to the water and sediment column, as follows similar research on cage effluent impacts on various sediment types [42]. Long-term monitoring of the offshore Royal Dag farm site in Ashdod from 2006-2008 also showed no impact to the surrounding water or sediment for a 500 m2, 350 tonne site [49].
Moreover, with the severe depletion of productivity in the Levant Basin in the immediate waters surrounding the offshore cages, we deduce that this ‘hot spot’ of metabolized wastes and excess fish food is being taken up by the adjacent consumers [50-51,15]. The super-low nutrient concentrations are typical artefacts of an ultraoligotrophic basin but are exacerbated by the apparent uptake of dissolved nutrients by primary producers [15]; these findings align with previous laboratory experiments of waste budgets and excretion rates of Sparus aurata [28,33]. These results also coincide with in situ observations from adjacent regional studies [43,42,52,15]. The in situ results and calculated bioavailable N per annum illustrate a small production mariculture farm with no significant impact to the water or sediment column at 700 t.
b. Effects of expanding the offshore industry
From large to small-scale farms, nearshore to offshore, significant changes to the water body and sediment characteristics are commonly cited as a limiting factor to the expansion of culture operations. These indicators or limits vary nationally, depending on the spatial-temporal scope of the industry and site characteristics and it is thus essential to employ a precautionary approach until short and long-term monitoring programmes are developed and baseline data on ecological impact is collected.
Countries have shifted a priori development via ‘trial and error’ in site selection and development, to utilizing more complex, advanced modelling techniques to determine the effect that culture will have, based on local conditions. The Scottish Development Agency [previously the Scottish Environmental Protection Agency] now enforces a ‘deposition-first’ approach through modelling, and strict environmental quality standards must be met before fish farm development [26], whilst other farms triage the chemical and biological oxygen demand of the water column [53]. Alternatively, the Norwegian government lists discharges of nutrient and organic material as goal 3 [after lice and diseases] of their national strategy [54]. In a normal trophic state, an increase in fish production would coincide with more metabolized cage wastes, bacterial activity, and greater local primary production [55]. A quantitative meta-analysis of publications by [56] reviewed the dissolved nutrient effect from aquaculture facilities. Their results indicated that ammonium had the greatest effect on the surrounding ecosystem, with nitrite-nitrate and phosphorus compounds following, and almost no effect from silicates. Ammonium and nitrites effluent as a result of fish farming was significant in comparison to multi-species farming, and the greatest descriptors to estimate effect.
Regarding the study’s small monoculture farm, the nutrient concentrations were greatest at five-fifteen meters below the sea surface, so it is likely that this input be taken up at the surface layers by primary producers. Per year, these producers will utilize 197,252 kg N. As the EMS system is significantly nutrient-limited, so the local waters of the Levant Basin can probably withstand the greater input of dissolved nutrients from mariculture. While proponents of DINTS (dilution is not the solution) may find fault with finding an acceptable input of nutrients for a given body of seawater, in the case of mariculture this metabolic waste is taken up by primary producers and may increase productivity levels in oligotrophic zones [15].
To this end, we compared the annual load of N to a consistent, long-term artificial input of solid phase waste from wastewater treatment at the Shafdan plant. This plant has operated from 1987 – 2017, and the outflow pipe was located 5 km offshore, at 37 m depth. At the mouth of the pipe [37 m depth] 2016 monitoring reported 6.25 µM ammonium [39], 7.3 times more than our highest observed value at the fish farms on the day. According to the National Action Plan of Israel, the annual loading of Total N at the mouth of the pipe was 2,982,696 kg N, which is 15 times greater than the annual loading of the fish farm [57]. Moreover, treated wastewater sludge contains a myriad of pesticides, detergents and heavy metals, whereas our input of artificial feed is a composition of nutrients, ash, fibre, and protein. In the most recent monitoring [2016] before the sludge outflow was redirected on land, the annual monitoring report states that ‘in all cases, Total N was lower than the value appearing in the water standard (1 mg/L)’ [39].
Conclusion
The in situ results and calculated bioavailable N per annum illustrate a small production mariculture farm with no significant impact to the water or sediment column at 700 t. Thus, the current monitoring scheme and production limit is amenable to the area. However, other allocated mariculture farms in Israel which are much larger in scale require stringent monitoring of nutrient effluent in consideration of different sediment types and changes in productivity levels.
This arbitrary standard was chosen by the Ministry of Environmental Protection in Israel, based on recommendations from the National Oceanic and Atmospheric Administration. The Shafdan sludge outflow was estimated to account for 85-98% of the pollution emissions from marine outfalls [39], and although it breaches environmental judgement, local fishermen did observe an increase in wild fish populations. The soluble effect was within the 1 mg/L threshold, and the effect of the incredibly high nutrient loading was expressed in the benthic zone (anoxic sediments immediately surrounding the discharge point). While we fully support the removal of the pipe, its comparison to the fish farm is pertinent to complement our assertions from the analysis.
The eastern Mediterranean Sea (EMS) is impacted by numerous factors [58], including unnaturally low nutrient concentrations and stratification of the water body in summer. Thus, studies that combine the concepts of assimilative capacity and the precautionary principle are needed more than ever. This research should be aligned with accepted, robust environmental quality standards from countries with a reputable status in offshore fish culture. Hence, further research on specific interactions and long-term monitoring is required as the industry expands its production to commercial scale. There is a significant paucity of fine-resolution data to support a monitoring programme for these proposed sites offshore, however, precautionary routine sampling of water and sediment must be implemented at all stages of production.
9. Acknowledgments
First and foremost, to Dr. Michael Krom for his unending support of this research from his initiation of the project, the collection, and full support during the analysis and drafting of this manuscript. The present work was funded by MERCI project of the University of Haifa and was conducted with great support during data collection and analysis from the staff at the Morris Kahn Marine Research Station My warm regards are extended specially to sampling team of the Michmoret Ruppin Academy (Professor Yair Suari and student team Tal Ben-Ezra, Shira Salingre, Matan Yuval, Liav Livne, and Sharon Confino) for their effort to collect water samples, sediment samples through SCUBA, and supportive analyses of water and sediment samples. Special thanks to the crew of the RV Medex and staff of EcoOcean for continually supporting our research. To Yossi Melchner and Ychiel Swimer of GiliOcean, Ltd. for answering the little questions about mariculture farms in Israel. To Tal Zvi Kedem of the Deep Med Laboratory of the University of Haifa for assistance with sediment sampling and analysis of TOC and grain size. To Eng. Noam Mozes, Alon Perlman, Dov Rosen for their detailed presentations on the projected mariculture space for Israel. To Drs Amir Neori and Muki Shpigel for their kind guidance and editing of this publication.
References
- UN General Assembly (2015) Transforming our world: the 2030 Agenda for Sustainable Development. UN General Assembly.
- Tveterås S, Asche F, Bellemare MF, Smith MD, Guttormsen AG et al. (2012) Fish is food-the FAO’s fish price index. PLoS One, 7(5), pe36731.
- Oyinlola MA, Reygondeau G, Wabnitz CC, Troell M and Cheung WW (2018) Global estimation of areas with suitable environmental conditions for mariculture species. PloS One, 13(1): pe0191086.
- Neumann B, Vafeidis AT, Zimmermann J and Nicholls RJ (2015) Future coastal population growth and exposure to sea-level rise and coastal flooding - a global assessment. PloS One, 10(3): pe0118571.
- FAO (2007) Cage Aquaculture: Regional reviews and Global Overview Technical Paper, 498.
- Campbell B, Pauly D (2013) Mariculture: a global analysis of production trends since 1950. Mar Pol, 1(39): 94-100.
- Gentry R, Froehlich HE, Grimm D, Kareiva P, Parke M, et al. (2017) Mapping the global potential for marine aquaculture. Nat Ecol Evol; 1: 1317–1324.
- Fernandez JD, Sanchez JP, Bayle SJT, Valle C and Dempster T (2008) Seasonal patterns and diets of wild fish assemblages associated with Mediterranean coastal fish farms. ICES J Mar Sci, 65: 1153–1160.
- Yokoyama H (2010) Monitoring, assessment and management of fish farm environments in Japan. Rev Aquac, 2(3), 154-165.
- Sanchez JP, Fernandez JD, Uglem I, Arechavala LP, Dempster T, et al. (2011) Coastal Fish Farms as Fish Aggregation Devices (FADs). CRC Press Taylor and Francis Group, 187-208.
- Ferreira JG, Saurel C, e Silva JL, Nunes JP and Vazquez F (2014) Modelling of interactions between inshore and offshore aquaculture. Aquaculture, 426: 154-164.
- Buck BH, Troell MF, Krause G, Angel D, Grote B et al. (2018) State of the art and challenges for offshore integrated multi-trophic aquaculture (IMTA). Front Mar Sci, 5: 165.
- Troell M, Rönnbäck P, Halling C, Kautsky N, Buschmann A (1999) Ecological engineering in aquaculture: use of seaweeds for removing nutrients from intensive aquaculture. J Appl Phycol, 11: 89-97.
- Mente E, Pierce GJ, Santos MB, and Neofitou C (2006) Effect of feed and feeding in the culture of salmonids on the marine aquatic environment: a synthesis for European aquaculture. Aquac Inter, 14(5), 499-522.
- Pitta P, Tsapakis M, Apostolaki E T, Tsagaraki T, Holmer M (2009) 'Ghost nutrients' from fish farms are transferred up the food web by phytoplankton grazers. Mar Ecol Prog Ser, 374: 1-6.
- Holmer M, Wildish D and Hargrave B (2005) Organic enrichment from marine finfish aquaculture and effects on sediment biogeochemical processes. In: Environmental effects of marine finfish aquaculture, 181-206.
- Holmer M (2010) Environmental issues of fish farming in offshore waters: perspectives, concerns and research needs. Aquacult Environ Interact, 1(1) : 57-70.
- Juergen V (2015) Prospects for Fisheries and Aquaculture.
- Aquarone MC, Adams S, Mifsu P (2009) LMEs and regional seas IV-7: the Mediterranean Sea.
- Antoine D, Morel A, and André JM (1995) Algal pigment distribution and primary production in the eastern Mediterranean as derived from coastal zone color scanner observations. J Geophys Res: Oceans, 100(C8): 16193-16209.
- Krom MD, Herut B and Mantoura RFC (2004), Nutrient budget for the eastern Mediterranean: Implications for phosphorus limitation. Limnol Oceanogr, 49: 1582–1592.
- Price CS and Morris JAJ (2013) Marine Cage Culture & The Environment. NOAA Tech Memo NOS NCCOS: 164-158.
- Bethoux JP and Copin MG (1986) Biological fixation of atmospheric nitrogen in the Mediterranean Sea. Limnol Oceanogr, 31: 1353–1358.
- Ribera d’Alcalà M, Civitarese G, Conversano F and Lavezza R (2003) Nutrient ratios and fluxes hint at overlooked processes in the Mediterranean Sea. J Geophys Res, Oceans, 108(C9).
- Brigolin D, Meccia VL, Venier C, Tomassetti P, Porrello S, Pastres R (2014) Modelling biogeochemical fluxes across a Mediterranean fish cage farm. Aquac Environ Interac, 5: 71-88.
- Cromey CJ, Nickell TD and Black, KD (2002) DEPOMOD — modelling the deposition and biological effects of waste solids from marine cage farms. Aquaculture, 214: 211-239.
- Jusup M, Ge?ek S, Legovi? T (2007) Impact of aquacultures on the marine ecosystem: Modelling benthic carbon loading over variable depth. Ecol Modell, 200: 459-466.
- Porter CB, Krom MD, Robbins MG, Brickell L and Davidson A (1987) Ammonia excretion and total N budget for gilthead seabream (Sparus aurata) and its effect on water quality conditions. Aquaculture, 66(3-4): 287-297.
- Almogi-Labin, A, Calvo, R, Elyashiv, R H, Amit, R, Harlavan, Y, Herut, B (2012) Sediment Characterization of the Israeli Mediterranean Shelf (10-100 m). Geological Survey Report, pp. 43.
- SEAL Autoanalyzer Methods: Seawater.
- Koroleff, F (1983) Determination of phosphorus. In: Methods of seawater analysis.
- Holm Hansen 0, Lorenzen CJ, Holmes RW, Strickland JDH (1965) Fluorometric determination of chlorophyll. ICES Journal of marine science, 30:3-15.
- Lupatsch I and Kissil GW (1998) Predicting aquaculture waste from gilthead seabream (Sparus aurata) culture using a nutritional approach. Aquat Liv Res, 11(4), 265-268.
- Uede T, Takeuchi T (2007) Postprandial changes in digesta, leftovers and feces, and their carbon and nitrogen contents in cultured red sea breams, Pagrus major. Aquac Sci 55: 409–415.
- Jansen HM, Reid GK, Bannister RJ, Husa H, Robinson SMC et al. (2016) Discrete water quality sampling at open-water aquaculture sites: limitations and strategies. Aquacult Environ Interact 8, 463–480.
- Neofitou N and Klaoudatos S (2008) Effect of fish farming on the water column nutrient concentration in a semi?enclosed gulf of the Eastern Mediterranean. Aquac Res, 39(5): 482-490.
- Pitta P, Apostolaki E T, Giannoulaki M and Karakassis I (2005) Mesoscale changes in the water column in response to fish farming zones in three coastal areas in the Eastern Mediterranean Sea. Estuar Coast Shelf Sci, 65: 501-512.
- Grossowicz M, Tchernov D and Gildor H (2017) A quantitative management tool reflecting impact of nutrient enrichment from mariculture in the Levantine Basin. Front Mar Sci, 4: 134.
- Ministry of Environmental Protection (2002) Monitoring Water Quality of the Mediterranean Sea.
- Ministry of Environmental Protection (2017) Setting background nutrient and chlorophyll-a levels and thresholds for the environmental protection of Israeli Mediterranean coastal waters: A statistical, experimental and modeling approach.
- Lavigne H, D'ortenzio F, d'Alcalà M R, Claustre H, Sauzède R, et al. (2015) On the vertical distribution of the chlorophyll a concentration in the Mediterranean Sea: a basin-scale and seasonal approach. Biogeosci, 12(16): 5021-5039.
- Karakassis I, Tsapakis M, Hatziyanni E, Papadopoulou KN and Plaiti W (2000) Impact of cage farming of fish on the seabed in three Mediterranean coastal areas. ICES J Mar Sci, 57: 1462–1471.
- Tsikopoulou I, Moraitis ML, Tsapakis M and Karakassis I (2018) Can intensive fish farming for 20 years induce changes in benthic ecosystems on a scale of waterbody? An assessment from Cephalonia bay (Ionian Sea). Environmental monitoring and assessment, 190(8), 469.
- Carroll ML, Cochrane S,Fieler R, Velvin R and White P (2003) Organic enrichment of sediments from salmon farming in Norway: Environmental factors, management practices and monitoring techniques. Aquaculture 226: 165-180.
- Morata T, Falco S, Gadea I, Sospedra J, Rodilla M (2015) Environmental effects of a marine fish farm of gilthead seabream (Sparus aurata) in the NW Mediterranean Sea on water column and sediment. Aquac Res, 46: 59–74.
- Brown, JR, Gowen, RJ and McLusky DM (1987) The effects of salmon farming on the benthos of a Scottish sea loch. J Exp Mar Biol Ecol, 109: 39-51.
- Hargrave BT, Duplisea DE, Pdeiffer E and Wildish DJ (1993) Seasonal changes in benthic fluxes of dissolved oxygen and ammonium associated with marine cultured Atlantic salmon. Mar Ecol Prog Ser, 96: 249-157.
- Weston DP (1990) Quantitative examination of macrobenthic community changes along an organic enrichment gradient. Mar Ecol Prog Ser, 61: 233-244.
- Israeli Oceanographic and Limnological Research Station 2006-2008 Monitoring of the fish farm site opposite Ashdod, the Royal Dag Fish Farm
- Cornelisen C (2013) Literature Review of Ecological Effects of Aquaculture. In: Chapter 5: Effects on Wild Fish. Ministry of Primary Industries, NZ.
- Uglem I, Karlsen Ø, Sanchez-Jerez P, Sæther BS (2014) Impacts of wild fishes attracted to open-cage salmonid farms in Norway. Aquac Environ Interact, 6 : 91-103.
- Karakassis I, Tsapakis M, Hatziyanni E, Pitta P (2001). Diel variation of nutrients and chlorophyll in sea bream and sea bass cages in the Mediterranean. Fresenius Environ Bull 10:278-283.
- OSPAR (2010) Quality Status Report 2010. OSPAR Commission, London: pp. 176.
- Anon (2009b) Strategy for an Environmentally Sustainable Norwegian Aquaculture Industry (in Norwegian), Oslo, Norway Ministry of Fisheries and Coastal Affairs. Pp. 34.
- Hixson SM (2014) Fish Nutrition and Current Issues in Aquaculture: The Balance in Providing Safe and Nutritious Seafood, in an Environmentally Sustainable Manner. J Aquac Res Development 5:234.
- Sarà G (2007) A meta-analysis on the ecological effects of aquaculture on the water column: dissolved nutrients. Mar Environ Res, 63(4) : 390-408.
- Ministry of Environmental Protection (2006) National Action Plan for the reduction of pollution of the Mediterranean Sea from land based sources.
- Katsanevakis S, Coll M, Piroddi C, Steenbeek J, Ben Rais Lasram et al. (2014) Invading the Mediterranean Sea: biodiversity patterns shaped by human activities. Front Mar Sci, 1: 32.